곤충과 해충
해충이라 부르는 벌레들
해충이라 하면 바로 떠오르는 것이 바퀴벌레일 것이다. 이는 주로 실내에서 발생해 유충과 성충 모두 집 안의 식품이나 생활쓰레기를 먹고 다량으로 발생한다. 이러한 음식 해충 가운데 쌀 등의 곡물에 생기는 해충을 저곡貯穀 해충이라 한다. 그 대표격이 어리쌀바구미이다. 이들 이외에 의류에 생기는 수시렁이 종류, 가구나 건축재에 생기는 흰개미나 나무좀, 책을 먹는 좀 종류, 나아가서는 인간이나 동물의 피를 빠는 모기, 파리, 벼룩, 이 등도 모두 해충이라 불린다. 사람에 따라서는 꽃의 꿀을 빠는 벌 종류나 바퀴벌레를 먹어 주는 거미조차도 불쾌하게 생각하는 해충의 일종일 것이다.
이처럼 '해충'이란 곤충을 포함한 절지동물 가운데, 인간의 생활권에 침입해 인간에게 직접 또는 간접적으로 신체적 고통이나 정신적 고통을 주는 무리를 가리키며, 인간이 일방적으로 마음대로 붙인 그들에게는 매우 불명예스러운 호칭이다. 사실 해충이라는 분류군은 생물학적으로는 존재하지 않는다. 저장 음식을 가해해는 해충 가운데 인간의 저장 음식 안에서 발견되는 거의 모든 곤충 종이 야외의 저장시설, 새의 둥지, 개미집, 또는 쥐 등의 설치류 보금자리 안, 흙 위의 익은 종자나 과실 위, 낙엽층, 나무껍질 아래, 또는 사체 위에서도 발견할 수 있기 때문이다. 대부분의 저장 음식을 가해하는 해충은 안정된 영양 기초와 번식 기초(식량)을 구하여 활발히 넓은 범위를 돌아다닌다. 그리고 그들이 영양 기초의 인공적인 퇴적 안 또는 위에서 대량 발생했을 때 처음으로 '저장 음식 해충'이라 불리는 것이다.
이 점에서 보아 해충은 인간이 만든 작은 환경에 생태적인 적응을 수행한 곤충으로, 저장 음식 곤충은 인간에 의한 식량의 저장이란 행위가 영속적으로 행해짐으로써 지속가능한 곤충이라 할 수 있고, 인위적 환경 없이는 생존할 수 없는 '재배식물'과 동종의 생물군이다. 이들은 대단히 문화적인 분류군이라 할 수 있다. 그들은 자연환경으로도 되돌아갈 수 있지만, 지금은 인간과의 공생(기생)의 길을 택하고 있다.
곤충의 생태
곤충의 종류는 현재 100만 종 이상이 알려져 있고, 추정되는 종 수는 300만~500만이라 한다. 덧붙여서 일본에서만 3만 몇 천 종의 곤충이 알려져 있다. 그들이 지구에 나타난 것은 약 4억 년 전으로, 400만 년 전에 출현한 인간보다도 훨씬 오랜 시대부터 영위해 온 동물로, 이 때문에 우리 인간은 지구에 나타났을 때부터 곤충과 깊은 관련을 가지고 있었다고 할 수 있다. 우리 고고학자가 연구대상으로 삼는 유적은 대략 몇천 만 년 전부터 최근까지 사이에 인간에 의하여 구축된 생활의 터전으로, 그곳에서는 곤충들의 유해가 사람과의 관련을 보여주는 상태로 발견된다.
곤충의 번식에 필요한 환경이나 그들이 요구하는 먹이는 종에 따라서 결정되어 있기 때문에, 곤충은 매우 소규모 주거 환경을 반영한다. 또한, 변온 동물이기 때문에 대부분이 기온에 예민히 반응한다. 곤충은 그들이 요구하는 기후 안에서는 진화적으로 안정되어 있으며, 기후나 환경의 변화 스트레스에 직면하면 유전적인 적응보다 오히려 적당한 장소로 이동해 먹이 선택을 넓히는 방법으로 적응을 하고 있기 때문이다. 그들은 제4기 동안 적어도 수백만 년 동안, 형태적·생태적·생리적으로도 거의 변화하지 않았다는 점, 그래서 환경사 복원에 매우 효과적인 지표라는 점이 잘 알려져 있다.
환경 지표로 유리한 곤층이 지닌 특성으로는,
1. 종류와 개체수가 많고, 서식 영역이 도처에 있다(자료의 풍부함, 광역성).
2. 특정 서식 환경을 보여준다(분포의 국소성).
3. 동정을 하기 쉽다(개성적 형태).
4. 수백만 년 동안 형태적·유전적 변화가 적다(유전적 안정성).
5. 온도 변화에 민감하고, 그에 따라 거주 영역을 옮긴다(고이동성).
등을 들 수 있다.
이와 같은 곤충이 지닌 특성에 기반해 해충류도 각각이 가해하는 대상물이나 생활역은 저마다 달라진다. 따라서 유적 안에서 발견되는 해충의 종류와 구성으로 당시 인간의 의식주를 알 수 있는 것이다.
해충화 징후군徴候群이란?
그럼 곤충은 어떻게 해서 해충화되는 것일까? 저장 음식 해충을 예로 그 메커니즘을 살펴보자.
저장 음식 해충을 비롯한 집해충과 인간의 일접한 관계가 성립하는 것은 앞에 기술했듯이 환경변화에 대응하는 곤충의 높은 생태적 적응력에 의한 것으로, 유전적인 변화 없이 인간이 만들어낸 다양한 환경에 적응하며 인간에 의존하는 생활형으로 그 생태를 변화시켰기 때문이다. 식물의 경우, 야생 식물의 인간에 의한 재배 행위가 시작되면 식물은 유전적 형질을 변화시키면서 인간에게 유용한 형질로 변화해 나아간다. 이들의 다양한 변화는 앞에 기술했듯이 '재배화 징후군'이라 하며, 종자가 튀어나가지 않는 점이나 종자가 커지는 점 등은 그 대표격이다. 해충의 경우, '공생이 아닌 '기생'이기에 인간 쪽에서는 장점이 전혀 없다. 이 의미에서도 인간이 정착 생활을 하게 되고, 음식을 저장하게 된 이후 약 1만 년 가까운 세월 동안 저장 음식 해충은 인간과의 일방적인 공생(기생)을 도모하기 위하여 진화를 이루어 온 곤충군이라 할 수 있겠다.
이 진화의 과정을 가령 '저장 음식 해충화'(이하 '해충화'), 그 저장 음식 해충화에 의하여 나타나는 생태적 특질을 '저장 음식 해충화 징후군'(이하 '해충화 징후군')이라 부르는 것이다.
그 대표적인 징후로는 아래의 4가지 점을 들 수 있다.
1. 비비상성非飛翔性)
2. 성충의 비섭식성非摂食性
3. 다화성多化性
4. 휴면성의 상실
날지 않는 성충
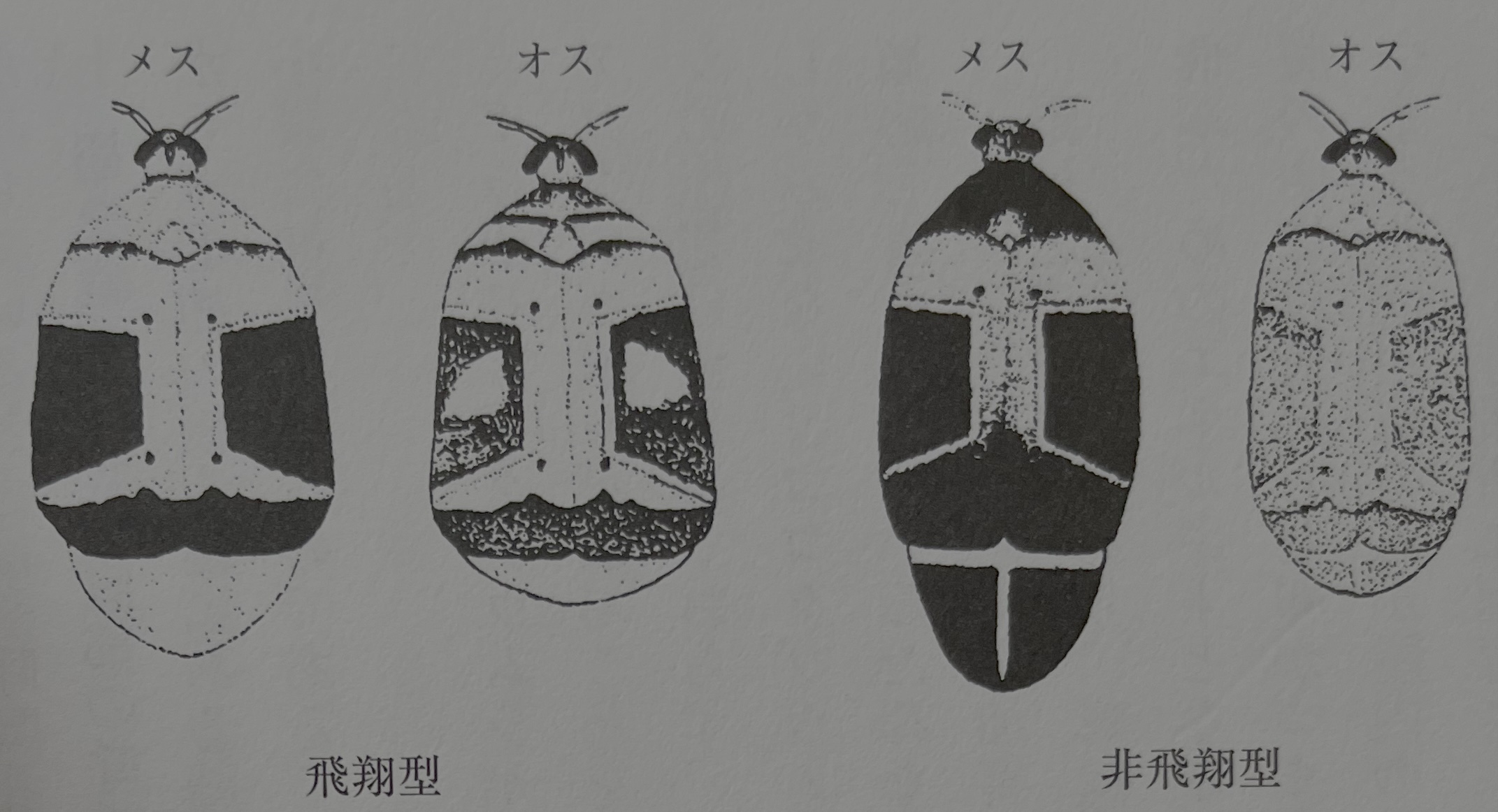
이를 보여주기 좋은 예로서 콩류를 가해하는 콩바구미과(Bruchidae)의 곤충이 있다(그림15). 해충 콩바구미류에는 그 생활형에 '야외형'과 '실내형' 2종이 있다. '야외형'이란 성충이 밭의 아직 익지 않은 콩의 꼬투리에 산란해 그 안에서 부화한 유충이 콩알의 안에 들어가, 그것을 먹으면서 콩의 성숙과 함께 자라는 것으로 일본에서 확인되는 해충 콩바구미류 가운데 누에콩바구미(Bruchus rufimanus BOHEMAN)과 완두콩바구미(Bruchus pisorum L.)가 이에 속한다. 이들에 반해 '실내형'은 팥바구미(Callosobruchus chinensis L.)와 네점콩바구미(Callosobruchus maculatus)가 그 좋은 예로, 유충운 완숙한 콩만으로 생활을 반복할 수 있다(梅谷 1987). 마찬가지로, 어리쌀바구미(Sitophilus zeamais MOTSCHULSKY)가 속하는 어리쌀바구미속 갑충의 자매종인 적갈색쌀바구미(Sitophilus granarius L.)와 일본의 벼바구미(Sitophilus oryzae L. 그림16)은 '실내형'으로, 그들은 완전히 인간이 만들어낸 인공적인 곡물 저장 시스템에 적응한 것이라 한다. 특히 적갈색쌀바구미는 오늘날까지 자연의 저장장에서는 발견되지 않는다고 한다. 이에 반해 어리쌀바구미는 실내형이지만 밀밭의 현장 가해도 기록되어 있어 야외형과 실내형의 절충형 생활형을 갖는다. 어리쌀바구미는 8월 무렵부터 월동 장소로 이동을 개시해 휴면한다. 그리고 이른봄에 꽃의 꿀을 빨고, 6월 무렵 창고에 밀 등이 운반되어 들어오면 저곡 장소에 침입한다. 적갈색쌀바구미나 벼바구미는 인간이 만들어낸 저장 음식의 영양 기초 안에서 1년 동안을 보내며, 기본적으로 날아다닐 수 없다. 그것은 저장장의 건조 환경에 비밀이 있다.


통상, 종자와 건조된 식물질 음식은 거의 수분을 포함하고 있지 않다. 따라서 곤충은 건조 음식 안의 탄수화물을 더 작은 분자로 분해(이화)하여 수분을 얻는다. 이와 같은 적응은 어리쌀바구미속을 포함한 대부분의 저곡 해충에서 볼 수 있다. 그들은 호흡에 의한 수분 손실을 최소한으로 억제하는, 즉 기체의 대사와 흐흡이 필요한 모든 행동을 중지함으로써 건조를 약화시켜 사막이나 곡물 저장소 같은 건조 환경 안에서도 살아갈 수 있게 된 것이다. 사실은 곤충이 날기 위해 필요한 산소는 기초대사의 10~100배나 높고, 많은 수분을 소비한다. 날기 위해서는 호흡구멍을 통한 호흡에 의한 가스의 증가가 필요하고, 날아다니고 있을 때는 호흡구멍이 완전히 열려 있다. 그러나 사막에 사는 대부분의 갑충 종이 윗날개를 유착시켜 복부의 호흡구멍 주위에 아랫날개의 방을 만들고, 그 안을 수증기로 채우면서 윗날개의 뒤쪽 끝에 있는 길쭉한 구멍으로 자연 호흡을 해서 호흡에 의한 물의 손실을 최소한으로 억제하는 것이다. 윗날개가 유착되어 있으면 날기 위한 아랫날개도 펼 수 없게 된다.
적갈색쌀바구미가 벼바구미에 비하여 건조에 대하여 강한 것은 적갈색쌀바구미가 나는 것을 포기했기 때문이고, 그들의 윗날개도 유착되어 있다. 벼바구미의 윗날개는 유착되어 있지 않고, 육체적으로는 가능하지만 그들은 거의 제한된 날기 행동만 보인다. 벼바구미가 비행 행동을 제한하고 있는 건 그들도 극학의 건조 조건에 적응하려고 하기 때문이다(Plarre 2010).
야외와 왕래하기
콩을 가해하는 네점콩바구미의 성충에게도 비상형과 비비상형이 있고, 형태적으로도 다른점을 볼 수 있다고 한다(그림17). 비상형은 날개가 길고, 날기 위한 에너지가 되는 지방량도 많은 데 반해 비비상형은 지방이 적은 대신에 배 안에는 성숙한 알이 가득 차 있어 우화 직후에도 곧바로 교미·산란을 행할 수 있다. 즉 비비상형 쪽이 실내의 콩 저장이라는 환경에서는 증식에 적합하다 할 수 있다.
그러나 보존된 콩이나 쌀은 무한하지 않다. 인간도 그것을 소비하고, 그들도 그것을 다 먹어 버리면 자손 등 남길 수 없다. 그래서 날아서 다른 콩 저장소나 야외의 콩밭으로 이동해야 한다.
그러나 생물이란 신기한 것으로, 날지 못했던 그들이 날 수 있게 되는 것이다. 거기에도 정확히 자연의 장치가 숨겨져 있다. 저장 콩 안에 산란을 반복하면, 유충의 밀도가 높아진다. 그러면 다수의 유충들의 신진대사에 의하여 나온 열이 생육에 영향을 주어, 발육이 늦은 개체가 발생한다. 이 몸이 약한 개체는 야외의 밭에서 자란 개체와 마찬가지로 오랜 시간에 걸쳐 천천히 성장한다. 그러면 날 수 있게 되는 것이다.
이 선조 되돌아감이라고도 할 만한 개체는 직접 다른 저장 콩으로 이동하든지, 그렇지 않으면 야외로 이동해 밭의 콩을 먹으며 연명하고, 그것에 알을 낳는다. 알을 포함한 밭의 콩은 수확, 저장되어 다시 그들의 번식 기회를 창설한다(梅谷 1987). 참으로 멋진 생태 적응이고, 생존 전략이기도 하다.
겨울철의 휴면에서 깨어난 어리쌀바구미가 꽃의 꿀을 빠는 모습이 목격되는데, 어리쌀바구미도 이 점에서 보아 야외형과 실내형 두 가지 형을 가지고 있다고 할 수 있다. 다만, 어리쌀바구미의 비상 범위는 전파원(농가의 곡식창고)에서 400미터를 넘지 않는다고 한다(吉田 외 1956). 또한 유럽이나 지중해 지역의 고고학 유적에서 발견되는 해충 중에서도 실내형으로 비상성을 갖지 않는 종이 4종류 정도 존재한다. 그중에서는 눈이 보이지 않는 종도 있고, 인간 및 가해물의 이동이 없으면 확산이 불가능하여 그 확산을 완전히 인간에게 의존하는 것도 있었다. 이런 점에서 보더라도, 어리쌀바구미속 갑충도 인위적인 영양 기초의 이동, 운반이 없으면 넓은 범위의 확산이 불가능한 해충이라 할 수 있다.
종자 안에서 생육하기
어리쌀바구미속 갑충이 번식하거나, 알곡에 적응하기 위한 또 하나의 중요한 것은 공생 박테리아를 소유하는 것이다. 이 박테리아는 평소는 성충 갑충의 세포질 안에 있는데, 난모세포를 통해 알이 형성될 때 다음 세대로 인계된다. 이 공생 박테리아는 기생주인 갑충과 영양소 사이의 효과적이며 효율적인 대사를 위한 상호작용이란 중요한 일을 행한다. 어리쌀바구미속 갑충은 추가 영양소가 없으면 곡물 위의 유충 성장에 극도로 지연을 일으키거나, 성충의 비상 능력을 잃어 버린다. 그러나 이 내부 공생자를 유지함으로써 어리쌀바구미속 갑충 각각의 종은 다양한 영양소와 번식 기초의 질에 거의 관계 없이 적응할 수 있는 것이다. 이 성질은 전분은 풍부하지만 편향된 영양소밖에 없는 알곡 위에서 발달하기 위한 중요한 요건의 하나가 된다.
어리쌀바구미속 갑충의 모든 종은 종자 안에서 번데기가 되어 성충이 되기까지 그곳을 떠나지 않는다. 이와 같은 적응은 인간에 의한 곡물 저장이 시작되기 전에 새나 설치류 같은 동물들이 종자를 채집해 그것을 장래 소비하기 위하여 안전한 장소에 보존하던 무렵으로 거슬러올라간다.
어리쌀바구미속 갑충이 인위적 환경(저장 종실)이나 자연 환경(야생 종실)이란 두 가지 이상의 환경에서 발달한 것은 신석기 시대에 인간에 의한 곡물의 재배와 비축이 시작되기 전에 이미 행해졌다고 생각된다. 조하리 씨는 비옥한 삼각지대 안의 나무 그늘 안의 잡초로서 야생 곡물이 자라난 졸참나무속 숲에서, 적갈색쌀바구미는 인간이 재배에 적응하기 전에 어느 설치류의 견과류 저장장에서 인간의 견과류 저장장으로 이동했다고 생각한다(Zohary 1969). 그 근거로는 맥류 등의 곡물 선조(야생)종의 이삭은 적갈색쌀바구미가 자라기에는 너무 작은 점, 이들이 지중해 주변부터 서쪽으로 확산된 것이 최초의 농경민들의 이동에 의한 것이란 점을 들고 있다.
이에 반하여 플라레 씨는 저장 곡물에 대한 가해는 서남아시아의 산악 산림 근처의 지역에서 일어났다고 상정한다(Plarre 2010). 여기에서는 재배 곡물과 함께 산악 산림에서 채집한 도토리도 같이 저장되어 있었다. 다만, 그 단계에서는 도토리도 곡물도 아직 다양한 채집 자원 가운데 하나의 구성요소에 지나지 않고, 종류나 구성도 안정적인 것이 아니며, 저장시설도 해충에게 그것들의 이용은 단속적으로 이루어질 수밖에 없었다. 그 때문에 당시는 사람이 재배한 저장 곡물 안에서만 번식하는 어리쌀바구미속의 한 무리는 발생하기 어려웠을 것이라 생각되고, 그 당시는 저장 곡물에 기생하던 갑충과 야외에 서식하는 갑충의 사이에 높은 레벨의 유전자 확산이 있었다고 생각한다. 또한, 저곡 해충인 바구미류는 적당한 온도이면 먹이가 없어도 몇 개월 살아갈 수 있다는 것이 잘 알려져 있으며, 어리쌀바구미속 갑충들은 일정 시기가 아니라 적당한 시기에 번식을 행하는 점이나 변화하는 먹이 자원에 적응할 수 있다는 성질을 이미 획득하고 있었다고 생각할 수 있다. 그것은 야외에서 그들의 본래적인 자연계 식용식물인 이엽시과(Dipterocarpaceae)와 졸참나무속이 일정하지 않은 시간에 익는 종자를 가지고 있었기 때문이다.

단명, 다산의 이유
'실내형' 콩바구미류에게는 교미나 산란을 위하여 성충이 되고나서부터 먹이를 필요로 하지 않는다는 공통된 성질이 있다(梅谷 1987). 이와 같은 성충은 우화했을 때는 이미 교미·산란할 수 있기까지 성장해야한다. 그러나 이 무리에게도 훌륭한 섭취 기관이 있고, 성충이 야외의 꽃에 와서 꽃가루를 먹고 있는 모습이 가끔 관찰된다. 또한 먹이를 먹이면 오래 산다는 실험 결과가 있다. 그때 교미를 시키지 않으면 더욱 수명이 늘어난다는 것이 알려져 있다. 이에 반해, 1년에 1번 발생하는 '야외형' 콩바구미류(섭식형)는 1년 동안의 생활사를 유충이 자라는 콩의 생육에 맞추어 조절하는 구조를 몸에 익히고 있다. 즉, 봄에 우화한 성충은 휴면하고 겨울을 나, 이듬해에 콩이 열매를 달기까지 산란을 기다릴 수 있는 것이다.
이에 반해 '실내형'은 1년에 몇 번이나 발생하는 다화성 성질이 그 기반이 된다. 즉, '실내형'의 콩바구미류는 완숙한 저장 콩의 위에서 콩을 섭취할 수 없기 때문에, 그 생명 연장의 전략을 영양분을 산란에 돌려서 그 발생 횟수를 늘린다는 생태로 변화시켜 적응한 것이라 생각할 수 있다. 이는 위에 적은 어리쌀바구미속 갑충과 똑같이 야외에서는 숙기가 정해지지 않은 1년 중 언제라도 종자를 볼 수 있는 녹두 야생종 등의 콩과식물을 기생주로 삼았던 선적응의 모습이라고 한다.
해충화의 야생 메커니즘
이상을 요약하면, 저곡 해충에게 볼 수 있는 기생주의 종자 알 속에서 완전한 발육을 행하는 점, 박테리아와 내공생하기, 그리고 건조 환경 속에서 수분의 손실을 막기 위한 비상 행동의 감소, 성충의 비섭식과 다화성 등은 인간에 의한 종자 저장의 개시 이전에 야생동물의 종자 저장에 대응하고 있던 무렵의 선적응이며, 결과적으로 이들 모두가 인간과 함께 존재하는 저곡 해충으로서 성공리에 생존하기 위해 유리하게 작용했다.
그리고 선사 시대에는 적어도 곡물이나 종자의 국소적인 교역(인위적 이동)의 가능성이 없으면 해충화하기 시작했던 집단의 확산은 불가능하고, 그들의 인위적 저장 음식에 대한 '적응 실험'은 실패했을 것이라 생각할 수 있다. 그러나 운 좋게도 인간들에 의한 종자의 저장과 공유, 그리고 저장 지역을 다계절에 걸쳐서 이용한다는 인간의 행위에 의하여, 음식의 전송에 따른 그들의 '보급 기회'가 만들어졌던 것이다(Plarre 2010).
유적 출토 곤충과 연구법
곤충 화석의 산출 상태
그럼 유적 안에서 곤충은 어떠한 상태로 발견되는 것일까? 곤충은 혐기성 퇴적물(저습지의 퇴적물 등) 안에서 식물 유존체 다음으로 가장 일반적으로 발견되는 동정 가능한 대형 화석이라 할 수 있다. 저습지 유적을 팠던 적이 있는 고고학자라면 누구나 풍뎅이나 비단별레 등의 무지개빛으로 빛나는 광택이 나는 껍질 조각을 발견하고 그 아름다움에 감동받은 경험이 있을 것이다. 또, 탄화된 똥이나 곡물 등의 물질 안에 포함된 곤충도 보존되는 경우가 있다. 곤충은 키틴질(다당고분자로 무척추동물의 몸 표면을 덮는 쿠틸라라 등의 주성분. 섬유상으로 강한 구조를 가짐)의 겉껍질을 가지기 때문에 습윤하든 건조하든 매몰 환경이 안정되기만 하면 유존 상태는 좋다. 특히 딱정벌레의 겉껍질은 영속성이 있고, 보통 파리는 번데기 상태로 남기 쉽다. 이 때문에 곤충은 고고학이나 환경사학 연구의 대상물이 되기 쉽고, 그 경우 저습지 퇴적물이 그 탐사의 주대상이 된다.
고고학적인 잔존 상황에는 아래의 네 가지 상태가 소개되어 있다(Panagiotakopulu 2000).
①습한 조건에서의 잔존 = 이탄층, 호수와 우물 등의 퇴적물
②탄화에 의한 잔존 = 탄화된 씨앗과 열매 및 그에 관련된 해충은 인위적 원인으로 일어나는 경우도 있고, 화산 분화 등으로도 일어남
③건조에 의한 잔존 = 매우 건조한 지대의 유적 특징
④광화(무기화 Mineralization)에 의한 잔존 = 방해석의 두터운 층에 의하여 덮힌 곤충의 빈 껍질
가장 오래된 적갈색쌀바구미의 화석은 토기 압흔이라고 한다. 이 토기 압흔은 ④의 광화에 의한 빈 껍질과 매우 비슷한 잔존 상황이다. 다만, 토기 안이나 토기 위의 압흔은 토기 소성에 따라 열에 의하여 생물체가 타면서 형성되는 것으로, 드물게 압흔 내부에 탄화된 곤충이 잔존하는 경우가 있다. 둘의 화학 변화 차이를 고려하면, 새롭게 '⑤토기 압흔'을 더할 수 있다. 이하 ①~③을 '생체 화석', ④와 ⑤를 '무기 화석'이라 부르고, 토기 압흔의 경우 단순히 '압흔'이라 표기한다. 일본의 고고학 유적에서 무기 화석의 좋은 예로는 고분에서 출토되는 철기의 녹으로 남은 파리 번데기 압흔을 들 수 있을 것이다.
단, 일본의 고고 유적에서는 주로 생체 화석, 그중에서도 곤충 유체를 가장 많이 출토하는 ①의 습한 조건에서 얻을 수 있는 것을 대상으로 연구가 진행되어 왔다.
일본의 곤충 고고학
일본 곤충 고고학의 1인자, 모리 유우이치森勇一씨에 의하면 고환경을 추정할 때의 지표가 되는 곤충은 다음 여섯 종류로 분류한다(森 2004). ①수역 환경(유수성流水性, 지수성止水性, 습지성湿地性), ②식생 환경(초원성, 방화성訪花性, 산림성), ③재배 및 농경(벼농사 해충, 밭농사 해충), ④오물 집적(식분성, 식시성食屍性), ⑤지표 환경(습윤, 모래땅 건조, 임상林床), ⑥기후 추정(한랭형, 온난형)이다. 이들은 곤충이 인위적으로 개변된 환경을 포함한 고환경의 복원 지표로서 유리한 것을 보여준다.
다만, 여기에서 볼 수 있는 곤충상은 자연환경을 축으로 하여, 인간이 관여한 환경(공간)도 '도시'나 '마을 산' 등 큰 생활환경의 구별에 머물러 '화장실'이나 '쓰레기장' 등의 일부를 제외하고 더욱 작은 주거환경을 보여주고 있지는 않다. 또한 해충으로 취급되는 것은 바구미를 제외하고 벼나 밭작물의 해충으로, 말하자면 야외형 곤충군뿐이다. 마찬가지로 곤충과 사람의 관계를 개략적으로 기술한 시야케 시게히코初宿成彦 씨 등의 저서에서도 화장실이나 쓰레기장 이외는 곤충 유체가 나타나는 주변 환경 이상으로는 좁혀지지 않는다(大阪市立自然史博物館 1996). 이러한 가운데 미야타케 요리오宮武頼夫 씨는 의식적으로 '실내 해충'이라는 분류군을 만들어 헤이죠우쿄平城京 터의 나가야오우長屋王 저택, 후지와라노 마로藤原麻呂 저택 출토의 쿠키 모양 탄화물 안에 들어간 바구미를 소개하고, 가루에 묻은 채 굳어졌을 가능성을 지적한다(宮武 1999). 게다가 헤이죠우쿄에서 발견된 머리대장가는납작벌레(Oryzaephilus surinamensis L.)나 거저리(Neatus picipes) 등의 저곡 해충도 소개한다.
그러나 대체로 유럽 등의 지역에 비하여 일본 곤충 고고학의 조사나 연구에서는 실내 해충에 관한 연구는 매우 저조하다고 할 수 있다. 저장 먹을거리 해충도 바구미나 톱가슴머리대장 류를 제외하고는 거의 알려져 있지 않다. 이는 그 분석대상이 저습지 유적의 이탄층 등에 편중되어 이루어진 데에 원인이 있는 듯하다.
서구의 곤충 고고학
유럽, 특히 영국에서는 환경 고고학 분야에서 곤충이 수행하는 역할은 매우 크다. 특히 인간이 개변한 환경에 곤충이 어떻게 적응했는지, 즉 도시에 있는 인간의 행동이나 생활환경을 복원하기 위하여 곤충이 그 증인이 되는 것이다. 예를 들면, 캐롯과 켄워드 둘은 앵글로색슨 시기의 요크 코파게이트에서 토양 속에서 출토된 750 이상의 시료에서 얻은 갑충을 중심으로 한 곤충류를 통계학적으로 해석해 실내상, 실내외의 부식물질, 고랑 등의 유기물이 많은 퇴적물상, 오수 웅덩이나 화장실상 등으로 분류했다(Carrott and Kenward 2001). 그리고 실내상에는 말린 풀이나 보릿짚 등의 건조 식물, 나무를 먹는 것, 사람에게 붙는 벼룩이나 이, 저장 먹을거리를 가해하는 것 등을 이 무리에 넣는다. 그리고 성격이 다른 퇴적물과 각각의 곤충 집단과의 사이에 어느 정도 상관이 있다는 것을 기술한다. 결과적으로, 도시라는 환경에 한하지만, 유적 출토의 곤충상부터, 실내 퇴적물이나 화장실의 퇴적물만이 아니라 고랑이나 불결한 가축의 사육장, 양모를 처리한 장소 등을 특정하는 것에 성공한다.
그러나 이와 같은 상황 가운데 아직 충분히 연구가 이루어지지 않은 것이 저장 먹을거리 해충이 어떻게 해서 고고학적 기록에 남은 것인지 하는 점이다. 이에 대하여 스미스와 켄워드 둘은 일정 모델을 제시했다. 영국 로마 시기의 퇴적물에서 자주 발견되는 저장 먹을거리 해충은 곤충상의 50~70%로, 많은 경우는 90%를 넘는다고 한다. 그 주요한 것은 적갈색쌀바구미, 톱가슴머리대장, 붉은양귀비톱가슴머리대장(Laemophloeus ferrugineus Steph)의 3종으로, 이들은 다량으로 출토된다. 이들 해충이 유적 퇴적물로서 출토되는 경로로 곡물의 수확 이후 가해된 곡물이 ①우연히 지붕재나 다른 것에 들어가 그들이 폐기되거나 무너진 경우, ②저장소에서 가해된 곡물이 우물이나 고랑에 버려지거나 소각된 경우, ③가해된 곡물이 다시 가축의 먹이나 맥주의 양조, 또는 인간의 식용으로 제공된 경우 등으로 나눌 수 있고, 최후의 인간 식사의 경우에 해충의 생체 화석이 화장실 유구의 토양 안에서 발견되는 것이 된다(Smith and Kenward 2011). 이것은 간략화된 상정 모델이지만, 어리쌀바구미 등의 해충이 유물이 되어 유적 퇴적물로 발견되기까지의 과정이 의외로 복잡했음을 예상하게 한다.
먹을 수 있었나? 저곡 해충
후쿠오카시에 있는 고대 영빈관迎賓館 터인 코우로칸鴻臚館 유적에서 최초로 화장실 유구가 발견되었을 때, 그 토양 안에서 어리쌀바구미의 생체 화석이 1점 검출되었다. 또한 이 화장실 유구의 인정 근거가 된 후지와라쿄藤原京의 화장실 유구 토양에서도 1점의 어리쌀바구미가 발견되었기에 마츠이 아키라松井章 씨는 이들을 사람이 먹었던 쌀 안에 뒤섞여 있던 것이라 해석했다. 확실히 어리쌀바구미를 먹고 인간의 배설물 속에서 소화되지 않고 정말로 나오는 것인지 하는 점에 대해서는 실제 실험이 행해져, 대부분 손상을 입지 않고서 배출된다는 것을 알 수 있었다(Buckland 1990). 여담이지만, 화장실 유구에서 다량으로 발견되는 박과 종자의 성인을 조사하기 위하여 똑같이 직접 먹고 배설물 속의 종자가 산출되는 상태를 관찰한다고 하는, 말하자면 몸을 걸고 실험을 행한 지인의 선생님이 있는 걸 떠올렸다. 알러지의 성인을 조사하기 위하여 기생충을 자신의 몸 안에서 키운 기생충학자도 있다. 동서양을 막론하고 학자라고 하는 사람은 얼마나 연구에 대해 진심이고 정열적인가를 느끼게 하는 에피소드이다. 용기 있는 실험을 행한 사람들에게 같은 연구자로서 마음으로부터 경의를 표하고 싶다.
경구 배설되었다고 생각되는 저곡 해충의 예는 이 일본 고대 화장실의 어리쌀바구미만이 아니다. 스코틀랜드의 바스딘バースデン에 있는 로마군 요새의 고랑에서는 적갈색쌀바구미가 들어간 밀이나 죽의 잔재가 보고되고 있고, 요크의 로마 시대 하수도 안의 저곡 해충의 덩어리는 인간의 배설물로부터 안으로 들어간 것이라 해석되고 있다. 앞의 캐롯과 켄워드 둘의 연구에서도 화장실 퇴적물의 곤충 집단에는 콩바구미 류가 포함되어 가해된 콩을 먹었던 결과라고 생각할 수 있다. 또한, 미야타케 씨가 제시한 어리쌀바구미가 들어간 쿠키는 해충에게 가해된 식품이라도 당시는 의외로 신경쓰지 않고 먹었는가 하는 생각이 들게 한다. 그렇다면 어리쌀바구미의 야요이 시대의 유일한 생체 화석의 예인 이케가미소네池上曽根 유적의 경우, 그들이 출토된 환호는 미야타케 씨도 지적했듯이 화장실로도 사용되었다고 하는 것이 된다.
그러나 해충이 배출한 요산이나 미세 식물(곰팡이나 이끼)이 늘어나면 그들이 독이 되어 인체나 가축에게 큰 영향을 준다는 것도 알려져 있다. 1432년 노팅엄의 예에 의하면, 토마스 샤프라는 인물이 어리쌀바구미속 갑충에 의하여 손상을 받은 맥아를 팔았고, 그에 의하여 사육된 돼지, 암탉, 닭이 죽어서 고소되었다는 사건이 일어난다.
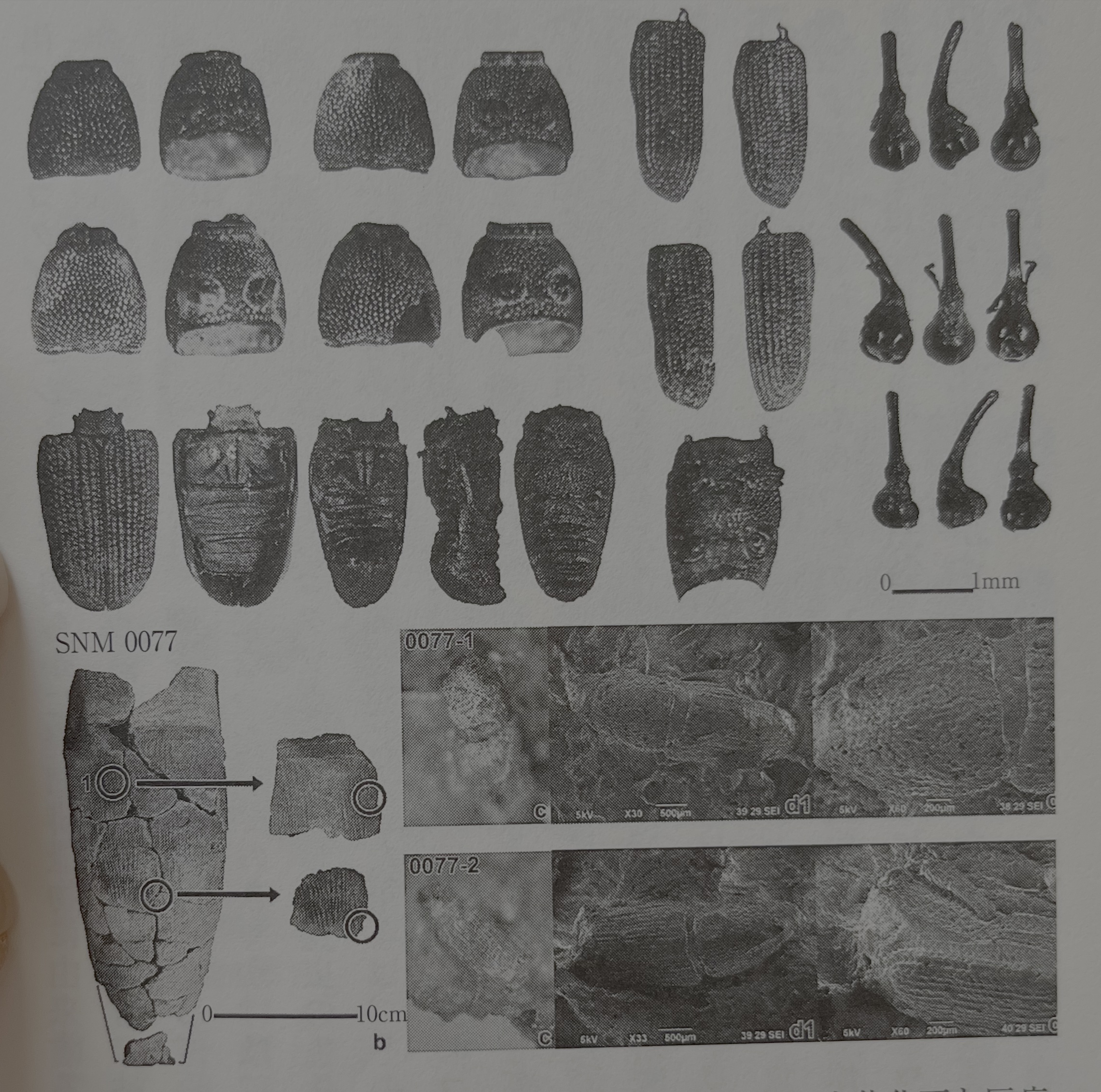
필자는 2003년에 발굴 조사된 코우로칸 북관의 화장실 유구 토양을 분석했을 때, 빵 상자 하나의 토양에서 다량의 박과나 가지과의 종자와 함께 132점의 어리쌀바구미의 생체 화석(부분)을 검출했다(그림19). 필자도 이 발견 전까지는 어리쌀바구미는 사람이 먹었던 것이라 의심하지 않았다. 그러나 지금은 상당히 발견 수가 많기 때문에 이 코우로칸 북관의 화장실 유구에서 발견된 어리쌀바구미의 생체 화석은 가해된 쌀과 함께 폐기된 것이 아닐까 추정된다. 그러나 앞의 '먹을 수 있다', 아니 '먹으면 독이 된다'는 논의는 가해의 정도를 잘 알 수 없기에 최종적인 결론을 내릴 수는 없다. 다만, 지금 우리들이 눈 앞에 두고 있는 토기 압흔으로 나오는 어리쌀바구미들은 명확히 경구 배설물에서 유래한 것이 아니다. 그럼 토기에서 발견되는 많은 어리쌀바구미들은 왜 토기의 점토 안에 들어간 것일까?

산나이마루야마三内丸山 유적에서 행한 검증
2012년부터 조사를 위해 아오모리시에 있는 저 유명한 특별 사적 산나이마루야마 유적을 방문하게 되었다. 그 계기는 한 논문의 사진 안에 산나이마루야마 유적에서 출토된 어리쌀바구미 같은 곤충의 생체 화석을 발견했기 때문이다. 어리쌀바구미에 남다른 친숙함을 느끼고 있던 필자는 그 사진을 보고 나는 듯이 산나이마루야마 유적으로 출발했다. 그곳에는 약 5000년 전의 어리쌀바구미속의 생체 화석이 실제로 발견되고 있었다(그림20 상단). 그 뒤, 몸이 더 크지만 확실히 어리쌀바구미라고 큐슈 대학의 모리모토 카츠라森本桂 씨가 동정해 주었다. 왜 이 본섬 최북단의 땅에 어리쌀바구미가 있는 것인지를 알고 싶어, 산나이마루야마 유적의 특별 연구에 응모해 토기 압흔에서 어리쌀바구미를 찾는 일을 했다. 결과적으로 5만 점 정도의 토기를 보고, 18점의 어리쌀바구미 압흔을 검출할 수 있었다(그림20 하단). 물론, 이들 압흔은 이 시점에서 열도 최북단 유적으로는 전국에서도 유일한 유적이고, 나아가서는 어리쌀바구미 이외에도 다수의 실내 해충이라 생각되는 곤충의 압흔을 검출할 수 있었기 때문에, 곤충 압흔의 성인에 대하여 깊이 생각하는 계기가 되었다.
국립역사민속박물관의 제1전시실에는 이 산나이마루야마 유적의 대형 건물과 그 앞마당 모습을 복원한 디오라마가 있다. 잘 만든 디오라마로, 이 건물 전면의 광장에서는 여성이나 아이들이 토기를 만들고, 그걸 굽는 모습도 표현되어 있다(그림21). 그러나 토기 압흔으로 다수 검출된 어리쌀바구미나 실내 해충은 토기 제작 장면이 이와 같은 실외가 아니었음을 호소하고 있다. 이 벌레들의 외침에 귀를 기울여 보면, 새로운 연구의 착상점, 즉 토기 압흔은 어떻게 형성되고, 무엇을 남기기 쉬운지 하는 의문과 압흔을 토기 제작의 기술이나 장소를 복원하는 소재로 사용할 수 없을까 하는 아이디어가 떠올랐다. 결론부터 말하자면, 토기 안에 뒤섞여 들어간 곤충이나 소동물은 실내 또는 그 주변에 서식하던 것이고, 토기가 제작된 장소도 그와 같은 곤충이나 소동물이 들어가기 쉬운 환경이었을 것이라 추정된다. 그때 토기 안의 생물체는 식물도 포함되고, 인간이 이용했던 것이나 인간과 관련이 깊은 생활환을 지닌 생물군이었을 가능성이 높다고 할 수 있다. 이 후보로서 식물의 경우는 야생 유용식물이나 재배식물이, 곤충의 경우는 실내 해충이 상정된다.

압흔과 생체 화석의 차이
엘리아스, 켄워드, 스미스 등의 연구에 의하면, 주거터의 상토에서 나온 곤충은 그 모두가 실내 해충이 아니라고 결론짓는다(Elias 2010, Kenward 1985, Smith 1996・2000). 그것은 벌레 자체의 침입도 물론이고, 사람이나 지붕에 둥지를 틀고 사는 쥐나 새 등에 의해서도 집 안에 운반되어 들어올 기회가 있기 때문이다. 그러나 인위적인 장벽을 뚫고 곤충이 토기 바탕흙에 들어가려면 여러 가지 샛길이 있지만, 집 안보다 조건은 어려워진다. 이와 같은 경우, 야외로부터 실내로, 실내에서 저장소로, 그곳들로부터 토기 바탕흙 안으로 이동해 가는 과정에서 곤충상의 종류와 수의 구성은 변화할 것이다. 즉, 야외에서 토기로 이어지는 도중에 곤충의 종류는 감소하고, 그것과는 반대로 1종류당 개체수나 실내 해충, 공생 해충의 비율은 증가할 것이다.
이 가설을 실제 자료로 검증해 보았다. 산나이마루야마 유적에서는 북쪽 골짜기 이탄층이나 제6차 조사구 등을 비롯해, 다섯 지점의 유적 토양에서 곤충 유체가 검출된다. 그중에는 풍뎅이 등의 똥벌레류나 밤바구미 등의 도토리 벌레, 노린재 등의 갑충 외에 초파리의 유충이나 번데기 등도 발견되어 그들을 먹으러 모인 반날개과나 풍뎅이붙이과, 딱정벌레과 등 식육, 식시성의 지표성 보행충이 다수 발견된다.
이상의 자료에서 토기 압흔을 더하면, 산나이마루야마 유적에서 발견되는 갑충, 소동물은 과와 목 수준에서 25종으로 나눌 수 있다. 이들을 그 생태로부터 자연에서 야외에 서식하는 '자연종'(일부 인간이 이용하는 식물에 식해를 가함), 오물이나 썩은 고기에 모이는 '위생종'(파리의 번데기나 유충), 인간이 사는 곳에도 서식하는 '공생종', 그리고 저장 먹을거리나 재료를 가해하는 '해충종'의 4종으로 분류했다.
이것을 앞의 가설과 비교해 보면, 종의 수는 압흔 자료의 쪽이 더 적어지는 것은 아니다. 오히려 해충의 종류가 증가하고, 야외의 자료와 거의 다름없는 수치를 나타냈다. 또한 1종류당 개체수는 가설과 달리 압흔 자료 쪽이 오히려 줄어든다. 이들은 토양 자료 안의 파리류의 번데기와 그들에 모이는 위생 해충의 많음에서 보이듯이, 해당 유적의 이탄층이 폐기물이나 배설물 등을 포함한 인위적 퇴적물로 완전히 자연 상태에서 이루어진 퇴적물은 아니기에 특정종의 수가 많고 종류도 한정되는 것에 기인한다. 그러나 이와 같은 인위적 퇴적물과 비교해도 위성종, 공생종, 해충종의 자연종에 대한 비율은 압흔 자료가 94%로 매우 높다. 해충종만으로 한정하면, 야외 퇴적물에서 가장 높은 30차 조사 자료가 21%인 데 반해 압흔 자료는 72%로 곤충상, 소동물상의 거의 3/4을 차지하고 있다. 유적 토양 자료는 자연종이 일정량 포함되어, 파리류 등의 위생종이 주체를 점하고 있는데, 압흔 자료에서는 해충종이 그들을 억제하고 80%라는 고율을 나타내고 있는 것이다(그림23).
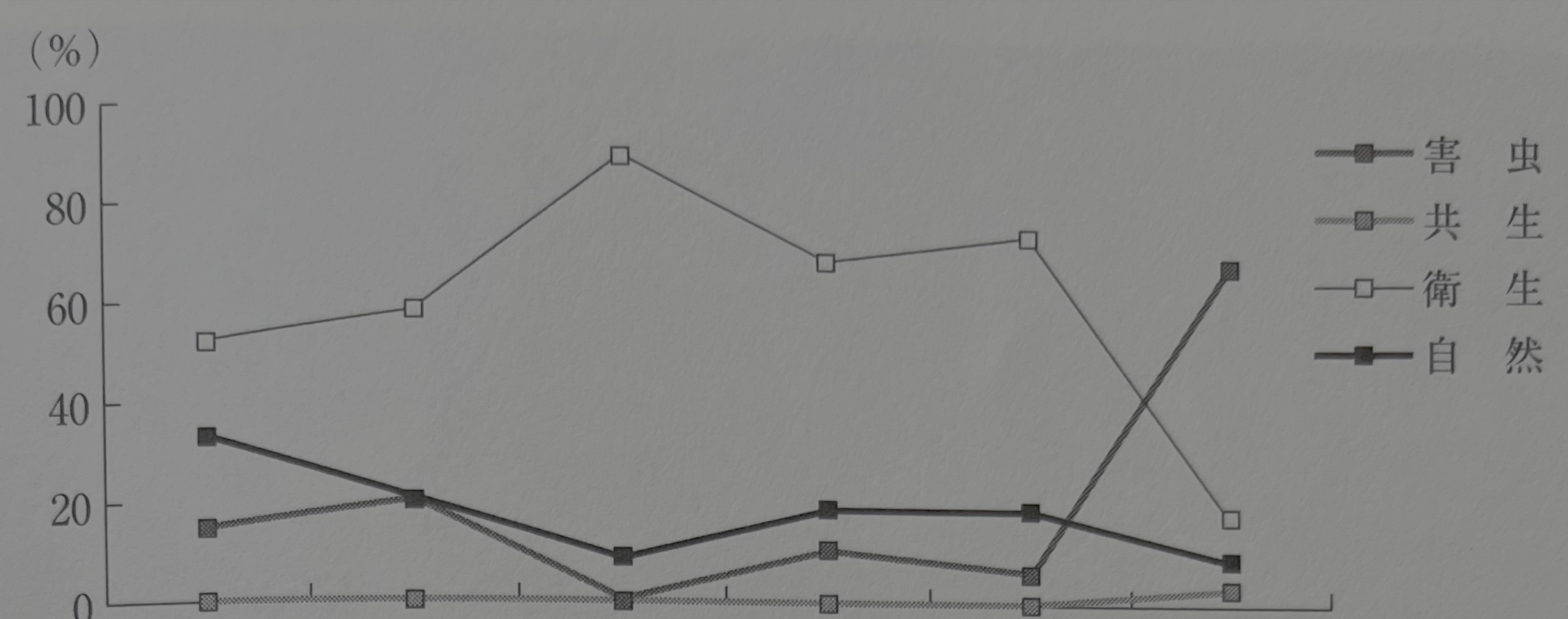
그림23 산나이마루야마 유적에서 자료별 곤충 조성비
전국의 벌레 압흔
2013년에 일본 전국 및 한국의 압흔 곤충을 집성했을 때의 수치에서는 어리쌀바구미속 갑충 이외의 곤충, 소동물 압흔은 총계로 40점이었다. 그 가운데 갑충목은 식물성 먹을거리(곡물)이나 목재를 가해하는 해충이 대부분으로, 어리쌀바구미 압흔을 더하면 개체수에서 전체의 80% 남짓을 점하고 있었다. 그외에 썩은 고기나 오물에 모이는 쌍시목(파리류)의 유충이나 번데기, 그들을 포식하는 풍뎅이붙이과나 송장벌레과의 갑충, 식분성 똥풍뎅이 등의 위생 곤충, 나아가서는 도토리나 밤 등에 기생하는 바구미과의 유충 등도 확인되어, 압흔 자료가 공생 곤충이나 해충을 매우 높은 비율로 포함하고 있는 것을 보여주었다. 그중에서도 실내형 해충이 비교적 높은 비율을 점하고 있었다.
이상을 종합하면, 똑같은 유적의 자료이면서도 토양 자료에 비하여 압흔 자료 쪽이 실내 해충이나 공생 해충의 비율이 높고, 토기 바탕흙 속에는 야외종의 뒤섞임이 적었다고 할 수 있다. 그리고 전국적인 경향도 이것을 뒷받침했다.
어리쌀바구미로 본 죠몬인의 삶
벼는 먹지 않았다
야마사키 스미오山崎純男 씨에 의하여 2005년에 처음으로 발견된 죠몬 토기의 압흔 어리쌀바구미는 구마모토현 오오야大矢 유적의 죠몬 시대 중기 말의 아타카식阿高式 토기의 현미 압흔, 구마모토현 이시노모토石の本 유적이나 타로우자코太郎迫 유적의 죠몬 시대 후기 중엽의 나락 압흔 등과 함께 벼의 죠몬 시대 도래설의 유력한 근거가 되어 왔다. 그 배경에는 연구자 사이에는 죠몬 후만기 농경론에 대한 막연한 신망이 있고, 그때까지도 죠몬 시대라고 주장되는 탄화 벼나 탄화 보리 등이 발견되고 있었기에, 형식과 시기가 확실한 토기의 압흔으로 벼가 검출되고, 게다가 그 해충인 쌀을 먹는 벌레까지 존재한다고 하면 누구나 벼의 전래 시기를 죠몬 시대 후기라도 틀림없다고 생각해 버린다. 나도 그 한 사람이었다. 오늘날에는 이들 벼 관련 압흔은 대부분 부정되고 있으며, 벼 및 벼농사의 수용 시기는 죠몬 시대 후기까지는 거슬러 올라가지 않는다는 것이 연구자 사이에서 대체로 일치를 보고 있다('벼는 언제 일본에 전해졌을까' 장을 참조). 사실은 어리쌀바구미에 대해서는 그 발견 당초부터 도토리나 밤 등의 전분질 식물 종자를 가해했다는 의견이 존재했다(安藤 2009).
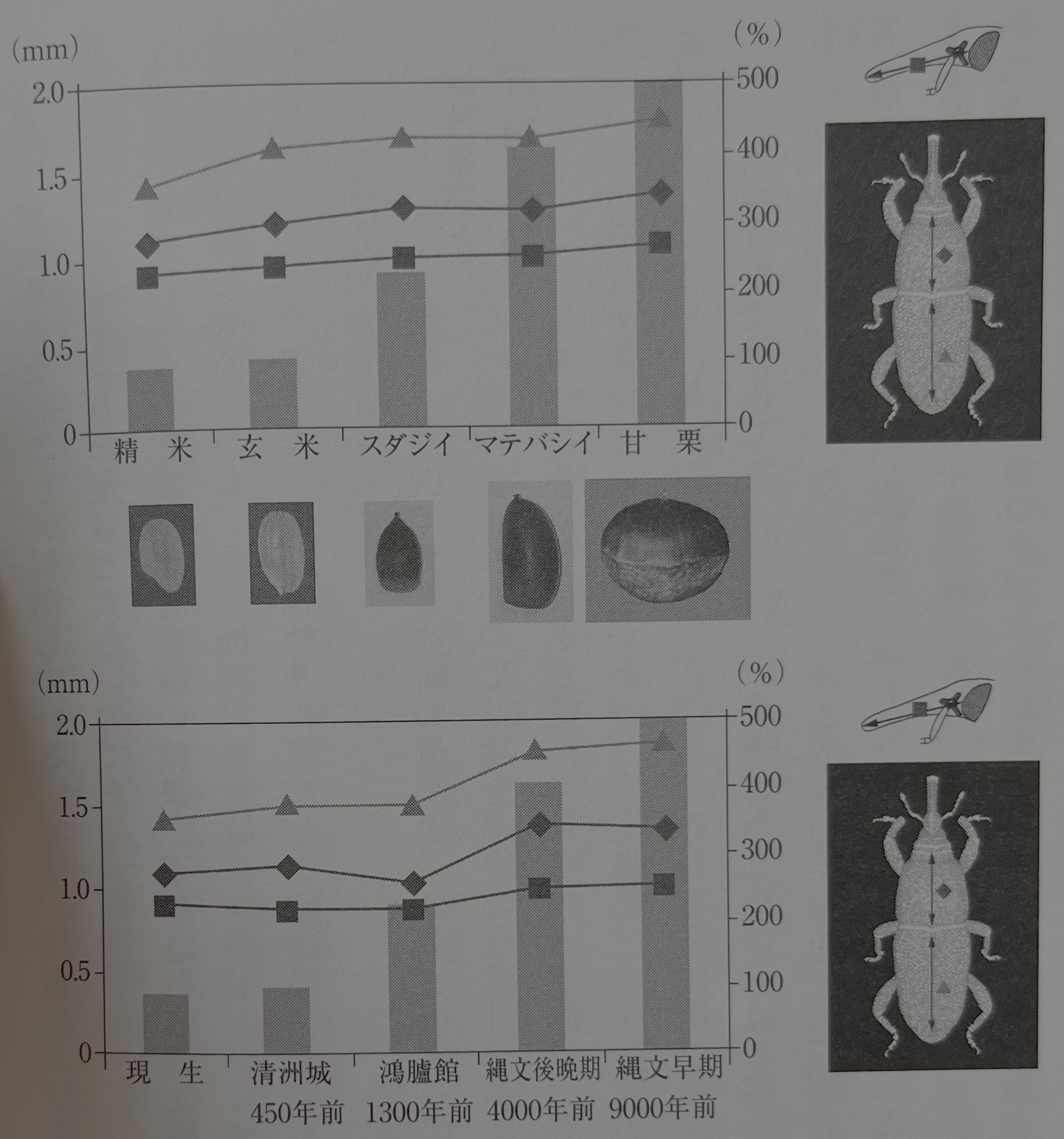
어리쌀바구미는 몸 길이 2.5~3.5밀리미리 흑갈색으로, 앞가슴등판이나 겉날개는 점각으로 덮여 있다. 그 유충은 쌀, 보리 등의 알곡 내부를 먹고 자라며, 알곡의 내부에서 번데기가 된다. 성충으로 월동하고, 1년 4번 정도 세대 교대를 한다. 이 어리쌀바구미가 포함되는 왕바구미 아과 갑충의 식성은 본래 외떡잎식물류의 살아 있는 나무에 적응해 성충이나 유충은 상당한 나무즙으로 채워진 외떡잎식물 조직 안(줄기나 뿌리)에서 생활한다. 따라서 어리쌀바구미를 포함한 어리쌀바구미족에게 쌍떡잎식물의 종자나 건조된 저곡물에 대한 적응은 소형화라는 형태 변화를 동반하고 있어, 2차적으로 일어났다고 생각된다. 자매종이라 생각되는 것으로 적갈색쌀바구미와 쌀바구미가 있다.
이들은 세계의 3대 저곡 해충이라 불리며, 그 기원과 확산에 관한 지금까지의 시나리오는 히말라야 남부의 숲 지대에 생식하고 있던 도토리 등의 종자에 적응했던 선조종이 인류에 의한 농경의 개시와 함께 채집 도토리와 재배 곡류가 함께 저장되는 환경 안에서 차츰 유전적 도태가 일어나, 곡류에 특화해 농경의 확산과 함께 구세계 각지로 확산했다는 것이다(Plarre 2010). 그러나 이들은 맥류 재배권인 서남아시아나 유럽에 주로 분포를 가진 적갈색쌀바구미에게는 적응할 수 있는 이론이지만, 벼농사 지대인 동아시아 지역의 경우는 고고학 자료가 부족해 이들 해충의 형성 과정은 전혀 알려져 있지 않았다. 동아시아의 대륙 쪽에서 어리쌀바구미족의 역사 자료로는 겨우 <이아爾雅>(기원전 5~기원전 2세기)에 보이는 어리쌀바구미라고 생각되는 저곡미 해충의 기록과 한나라 대의 마왕퇴馬王堆 한묘 1호묘에서 검출된 쌀바구미의 생체 화석뿐이었다. 일본에서는 오사카부 이케가미소네池上曽根 유적(야요이 시대 중기), 나라현 후지와라쿄 좌경左京 칠조일방서북평七条一坊西北坪(7세기), 같은 후지와라쿄 나가야長屋 왕 저택(8세기), 후쿠오카현 코우로칸鴻臚館(8세기), 아이치현 키요스清洲 성하정城下町 유적(16세기 말~17세기 초두) 등에서 생체 화석이 별견되었는데, 야마사키 씨에 의한 죠몬 시대의 어리쌀바구미 압흔의 발견까지는 벼농사 도래기 이전의 상황을 보여주는 예는 전무했다고 할 수 있다. 이 의미에서도 죠 몬 시대의 후만기 압흔 어리쌀바구미는 동아시아에서 가장 오래된 예였다.
곤란한 대발견
필자 등은 2010년 2월에 가고시마현 타네가시마種子島를 방문해, 죠몬 시대 초창기~조기에 걸친 토기 압흔 조사를 실시했다. 가고시마 대학의 나카무라 나오코中村直子 씨에 이끌려 죠몬 시대 초창기 먹을거리의 흔적을 압흔으로 찾고자 했던 것이다. 우리는 여기에서 의외의, 참으로 생각 외의 발견을 하게 된다.
초창기 오니가노鬼ヶ野 유적의 압흔 조사의 결과가 부진했기에 니시노오모테시西之表市 교육위원회의 오키타 준이치로沖田純一郎 씨에게 죠몬 시대 조기의 산본마츠三本松 유적에서 출토된 토기를 보여 드렸다. 조사도 종반에 접어들어 돌아갈까 생각했던 즈음, 초대 연구 조수인 센바 야스코仙波靖子 씨가 머뭇거리면서 "이 유적, 조기이죠..."라고 불안하게 물어보았다. "당연하지"라고 답하자, "나왔습니다"라고 한다. "뭐가?"라고 묻자, 토기를 내밀며 "어리쌀바구미입니다"라고 작은 목소리로 중얼거렸다. 반신반의하여 실물현미경으로 들여다보자, 렌즈 아래에 확실히 낯익은 점각 줄이 있는 겉날개나 흉부가 나타나 한순간 눈을 의심했다. 토기는 틀림없이 죠몬 시대 조기의 요시다식 원통 토기의 몸통 조각으로, 설마 하고 생각했지만 돌아오기 직전에 다른 정리사무소에 두었던 토기 조각 안에서 또 1점의 어리쌀바구미 압흔을 검출할 수 있어 죠몬 시대 조기의 것임이 틀림없다고 확신했다.
연도가 끝나는 4월에 조속히 니시노오모테시로 향했다. 2대째 연구 조수인 마나베 아야真邊彩 씨와 남은 토기를 이 잡듯이 조사해 보기로 했다. "아무것도 나오지 않네"라고 체념하고 있을 무렵, 둘이 동시에 '나왔다!"라고 외쳤다. 그리고 그 뒤 1시간 사이에 총계로 5점의 어리쌀바구미 압흔이 나왔다(그림24). 어리쌀바구미의 압흔만이....... 밖은 비가 내리고 있었다. 그 시간이 지나자 이제 1점도 발견할 수 없었다. 학자도 징크스나 미신을 믿는다. 특히 귀중한 발견을 했을 때는 우연(필연)이라 생각하면서도 무언가 자신의 힘만이 아님을 느끼고 그에 의지하려 한다. 그만큼, 뛰어난 신이 내린 발견의 순간이었다. 이 성과는 나중에 미국의 과학 저널 PLOS ONE에 "세계에서 가장 오래된 저장 먹을거리 해충"이란 제목으로 세계에 발신되었다.

계속 늘어나는 압흔 어리쌀바구미
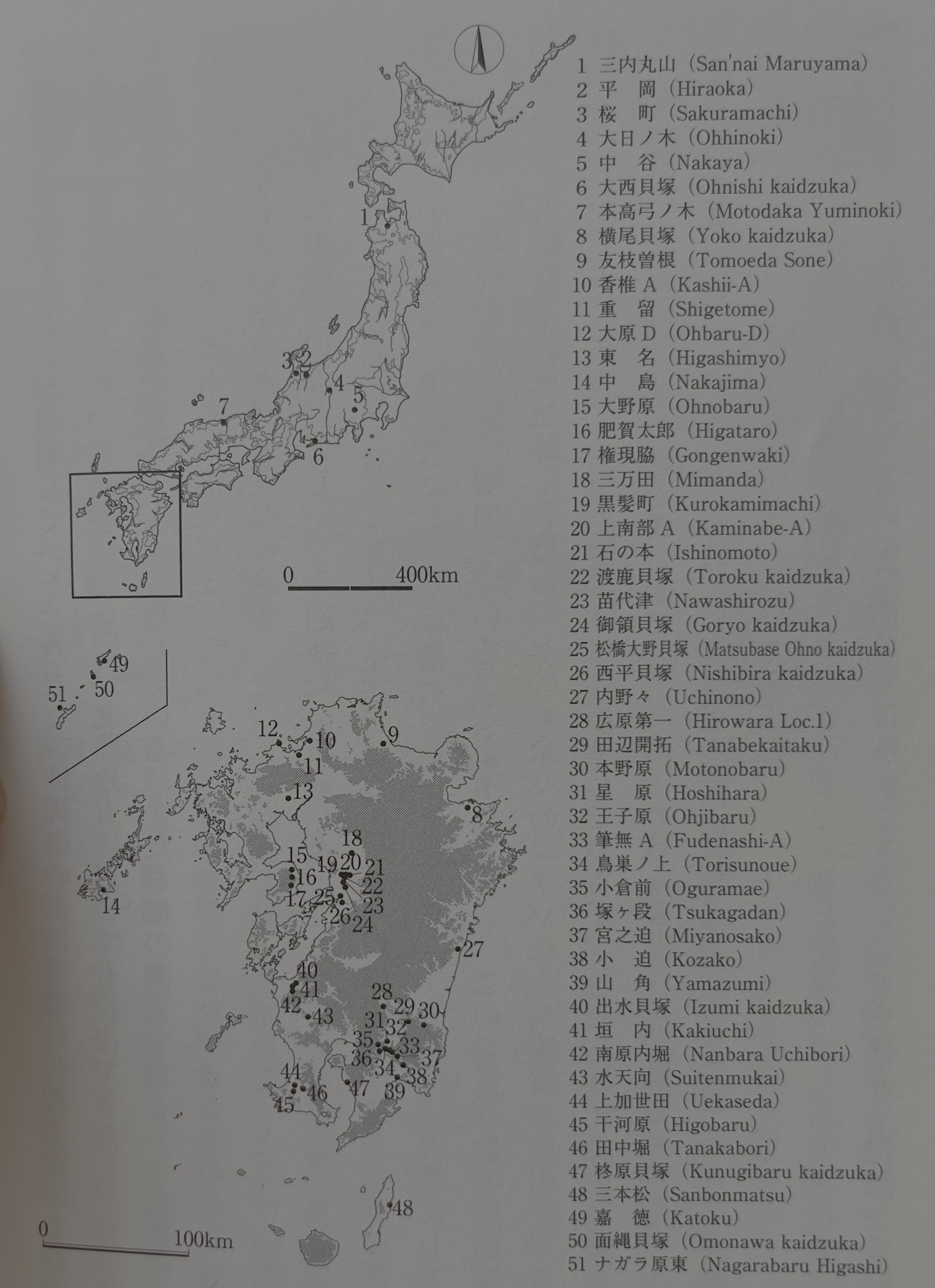
2011년의 시점에서 죠몬 시대의 어리쌀바구미 압흔은 17유적 44예였는데(小畑 2011), 그 뒤 필자 등의 조사에 의하여 1년 사이에 34유적 112예까지 증가했다(小畑 2012). 이 시점에서 북쪽은 아오모리현, 남쪽은 오키나와현까지 거의 전국적으로 분포한다는 것이 판명되었다. 다만, 오키나와현의 예는 고분 시대 상당 시기의 것이었다. 그 뒤 필자가 행한 조사에서, 호쿠리쿠北陸 지방에서도 검출할 수 있었고 아마미奄美 제도, 고토우五島 열도 등의 도서부 유적에서도 검출된다. 또한 다른 조사자에 의해서도 여러 예의 보고가 이루어진다. 이들의 성과에 의하여 서서히나마 어리쌀바구미의 분포역이나 출토되는 시기도 그 공백이 매워져 갔다. 다만, 현재로서는 큐슈 지방, 특히 남큐슈나 남서 제도의 사례 증가가 두드러진다. 이번 새롭게 집성한 바, 51유적 325점으로 증가했다(그림25). 또한 그림 26에서 보듯이, 압흔 어리쌀바구미의 검출수는 그야말로 승승장구하고 있다. 이 수치는 유적 수에서 3년 전의 3배, 2년 전의 1.5배, 압흔 수에서 3년 전의 7배, 2년 전의 약 3배로 증가한 것을 보여준다. 이 발견 개수가 극적으로 증가한 배경에는 미야자키현 모토노바루 유적에서 170점 남짓의 어리쌀바구미 압흔이 검출된 것이 크게 관련되어 있는데, 잘 보면 유적 수는 그다지 증가하지 않았고 오히려 연엑스선이나 엑스선 CT에 의한 잠재 압흔의 검출('압흔법이 밝힌 것'이란 장을 참조)이 유적 수나 개수의 일부를 증가시킨 요인이 된다. 지금으로서는 "일본 전국의 죠몬 유적 어디에서도 검출할 수 있다"고 반은 진심으로 허충을 떨고 있을 정도이다.
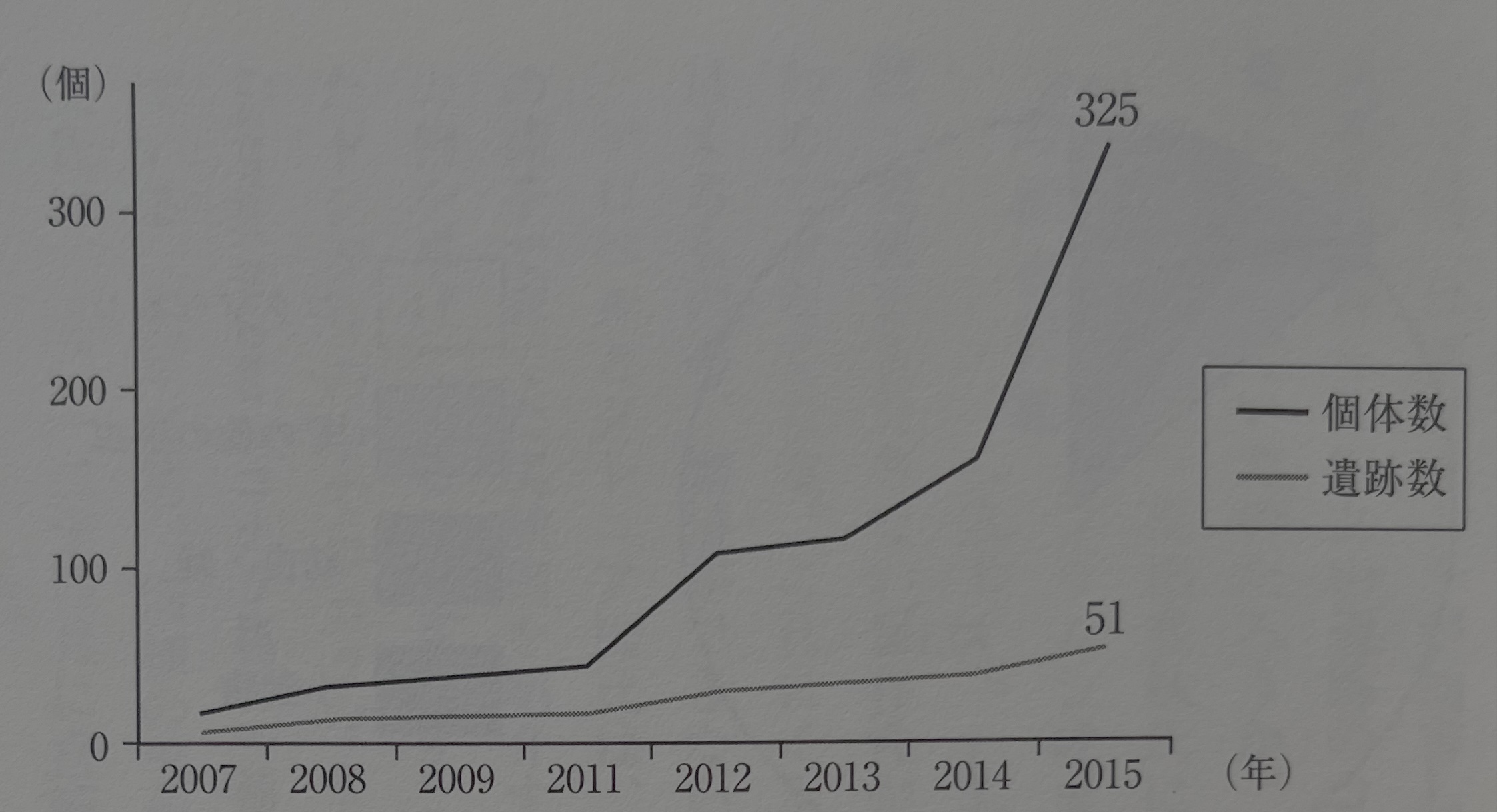
이러한 넓은 시대와 지역에서의 발견은 최근까지 벼와 관련되어 왔던 죠몬 시대의 어리쌀바구미가 벼농사의 도래라는 전파론으로는 해석할 수 없다는 것을 웅변하고 있다. 아래에서 보는, 어리쌀바구미의 의미에 관한 새로운 해석은 벼를 먹는 현대의 어리쌀바구미의 진화나 성립 과정에 대하여 새로운 빛을 비추는 듯하다.
어리쌀바구미가 의미하는 것
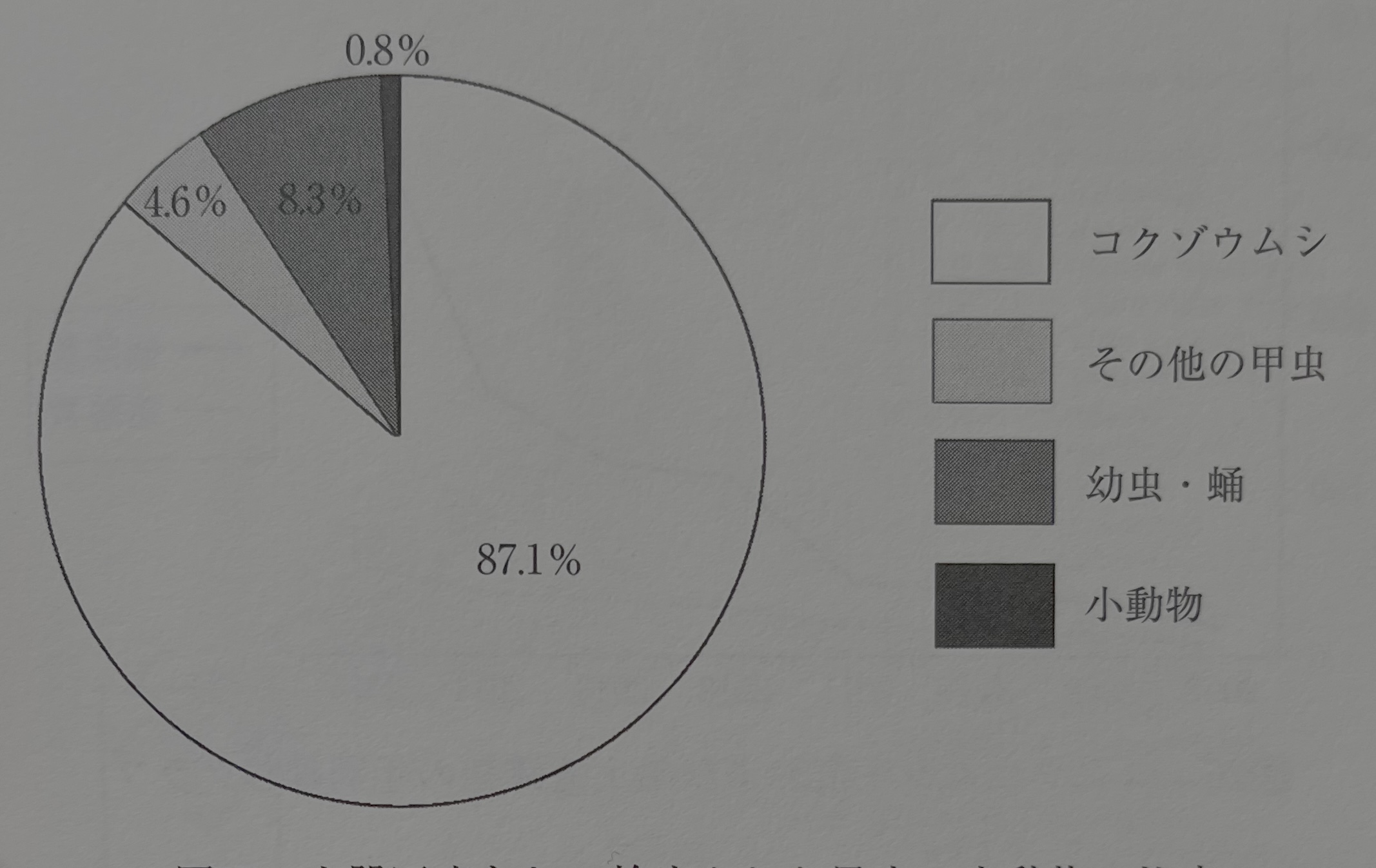
어리쌀바구미 압흔은 앞에 기술했듯이, 2015년 6월 현재 325점이고, 이때까지 발견된 그외의 곤충, 소동물의 압흔 총수는 51점이었다. 따라서 어리쌀바구미는 곤충, 소동물 압흔 수의 약 90%를 차지해 점유율로는 이상하게 많다고 할 수 있다. 사실은 압흔으로는 많이 나올 것 같은 개미의 압흔은 1점도 검출되지 않는다(그림27).
나아가 놀랄 만한 것은 하나의 토기(조각)에서 복수 개체의 어리쌀바구미 압흔이 검출되는 예가 있다는 점이다. 산나이마루야마 유적에서는 원통하층식 토기 1개체에 2점, 원통상층식 토기에서 3점, 손바닥 크기의 원통상층식 토기의 파편에서 3점과, 1개체의 토기에서 복수의 어리쌀바구미 압흔이 생겨 있었다. 또한 가고시마현 소오시曽於市 오구라마에小倉前 유적에서는 돌대문突帯文 토기 시기의 단지형 토기의 주둥이 부분 조각에서 3점, 같은 시 츠카가단塚ヶ段 유적에서는 얕은 주발형 토기의 주둥이 부분 조각에서 6점의 어리쌀바구미 압흔이 생겨 있었다.
이 유적 수나 검출 개체수에서 보이는 높은 검출률, 그리고 곤충 압흔 중에서 90%라는 높은 비율은 어리쌀바구미가 토기 제작 환경이었던 실내에 많이 서식하고 있었단 것을 보여준다. 비슷한 현상은 야외에서 그들을 채집해 와서, 의도적으로 토기에 넣지 않는 한 일어나지 않는다. 이와 같은 행위는 완전히 있을 수 없는 일은 아니지만, 어리쌀바구미 압흔은 죠몬인들의 집에 다량으로 발생한 해충으로, 그 가해 대상이 죠몬인들의 집이나 마을 안에 있던 것을 의미한다고 생각한다.
그럼, 어리쌀바구미가 가해한 저장 먹을거리는 무엇일까? 벼 전파의 경로나 시대에서 멀리 떨어진 산본마츠 유적이나 산나이마루야마 유적의 어리쌀바구미들은 대체 무엇을 먹어서 토기 압흔으로 남을 정도로 다량으로 발생했던 것일까?
어리쌀바구미의 가해 대상
하라다 토요아키原田豊秋 씨에 의한 어리쌀바구미가 가해하는 먹을거리의 목록에 의하면, 어리쌀바구미 성충이 먹고 생존할 수 있는 것은 37과 96종에나 이른다(原田 1971). 그러나 유충의 먹을거리는 유충 1세대를 지탱할 만한 용적과 부패, 변질을 일으키지 않는 보존성이 있는 것이 요건으로, 먹을거리의 종류는 11과 30종으로 매우 적어진다. 이것에 벼를 더하면 31종이 되지만, 성충의 먹을거리와 비교하면 먹기는 하지만 번식을 할 수 없는 것이 60종 이상 존재하게 된다.
들로벨과 그르니에 둘은 어리쌀바구미와 적갈색쌀바구미를 졸참나무속과 밤의 종자에서 키우는 실험 예를 소개하며 도토리 안에서 발견된 어리쌀바구미속 갑충의 예를 든다(Delobel and Grenier 1993). 주버트 씨는 남아프리카 너도밤나무의 깨진 도토리 안에서 바구미를 발견하고(Joubert 1966), 밀스 씨는 캔자스의 인도산 참나무(Quercus incana, 졸참나무속의 일종, 일본식 이름 없음)의 깨진 종자 안에서 어리쌀바구미를 발견한다(Mils 1989). 똑같은 사례는 근처에서도 일어났다. 도토리 연구의 제1인자 이와나가 테츠오岩永哲夫 씨는 2012년 4월에 미야자키현 사이토시西都市의 미나미카타南方 신사에서 채집한 모밀잣밤나무, 가시나무, 종가시나무의 안에서 어리쌀바구미 몇 마리를 발견했다. 필자는 그 가운데 한 마리를 나누어 받아서 쌀로 사육하던 중에 실수로 으깨 버렸다. 낙담하고 있던 차에 그녀가 알을 낳았던 듯하고, 그 6월 쌀 안에서 36마리의 차세대가 탄생했다는 전말이 있다(그림28).
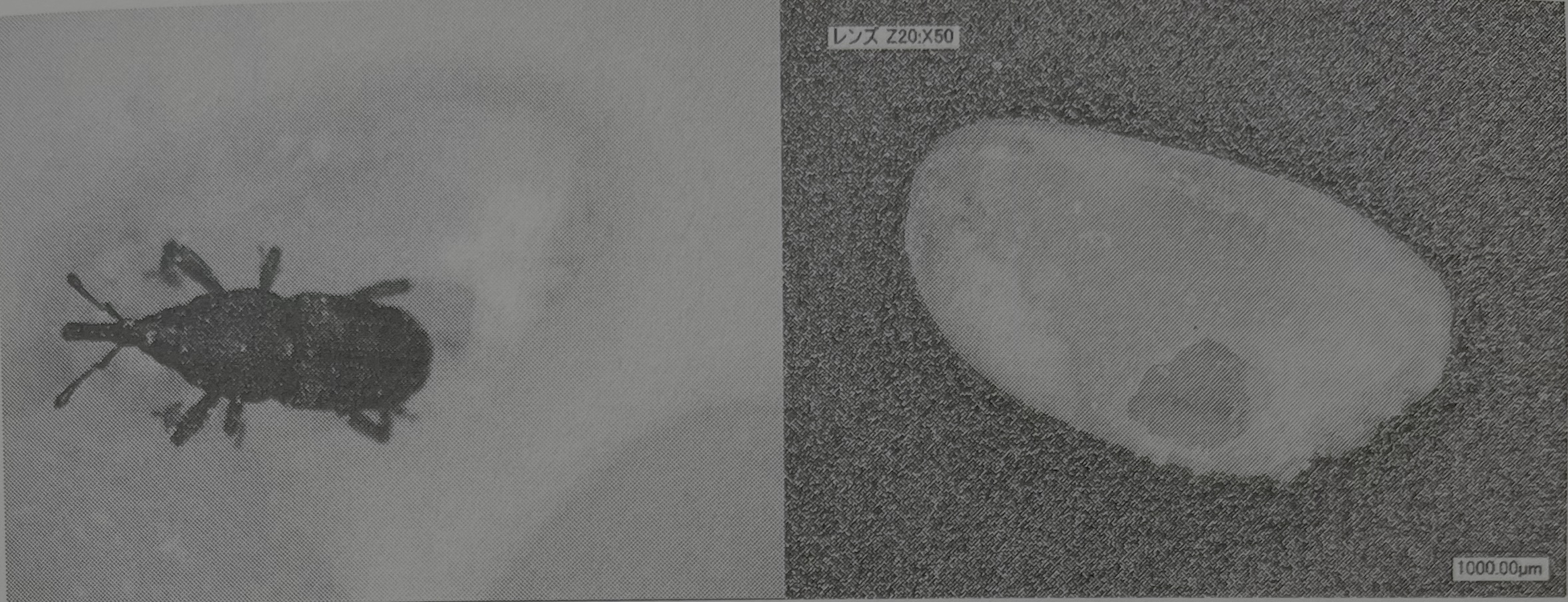
세계에서 최초로 적갈색쌀바구미를 졸참나무속의 종자와 밤에서 키우는 일에 성공한 것은 자허 씨였고(Zacher 1937), 들로벨 씨 등의 생육 실험에서도 밤과 도토리(졸참나무속) 종자에 대한 높은 적응이 보고된다. 필자도 이들의 보고서를 읽고, 도토리의 종류에 따라서 어떻게 생육 상황이 다른지를 알고 싶어서, 평소 어리쌀바구미 등의 저곡 해충을 죽이는 연구를 하고 있는 식품종합연구소의 미야노시타 아키히로宮ノ下明大 씨에게 견과류에서 생육하는 실험을 의뢰했다. 그 결과, 우화의 시간이나 수는 종류마다 다르지만, 실험에 사용한 밤, 구실잣밤나무, 가시나무, 모밀잣밤나무 모두에서 산란해 견과를 먹고 성충이 되었다. 다만, 겉껍질에 상처가 없는 견과에는 산란할 수 없고, 식용으로도 할 수 없다는 것이 판명되었다. 한편, 깨지거나 구멍이 있는 견과의 경우, 과육만 있는 견과의 경우와 거의 다름없는 우화 수임을 알 수 있었다. 이 실험의 결과로부터, 죠몬 시대에 실내에서 겉껍질을 벗긴 상태로 견과류가 건조되어 저장된 경우, 또는 겉껄집이 붙어 있는 상태로도 건조로 갈라짐이 생긴 견과가 있었던 경우에는 어리쌀바구미가 발생할 가능성은 충분한데, 어리쌀바구미가 번식하기 위해서는 몇 개월 단위로 그들이 보관될 필요가 있다고 하는 결론을 얻을 수 있었다(宮ノ下 외 2010).
본래 참나무과 식물의 종자에 적응해 진화해 왔던 곤충군이기에 밤이나 졸참나무속의 종자를 영양이나 번식의 기반으로 삼는 일은 지극히 당연할 것이다. 다만 이삭 달린 벼의 경우와 마찬가지로 겉껍질이 있으면 번식할 수 없는 점은 견과류의 저장법이라는 의미에서 중요하다.

커다랐던 죠몬 어리쌀바구미
어리쌀바구미는 통상 3~4밀리미터이다. 크기의 변이는 있지만, 거의 이 정도의 크기이다. 성충의 몸길이는 발생 시기로도 달라진다. 여름 시기 발생한 것은 봄철이나 가을철 발생한 것보다 소형으로, 평균으로는 몸길이 0.2 밀리미터 정도 차이가 있다고 한다(原田 1971). 또한 문헌에 따라서 그 계측치에 큰 차이를 볼 수 있다.
하라다 토요아키 씨의 문헌은 평균 3.8~4.0밀리미터라고 표기되어 있는데(原田 1971), 야스토미 카즈오安富和男 씨 등은 2.3~3.5밀리미터(평균 2.8밀리미터)이다(安富·梅谷 2000).
어리쌀바구미는 애벌레 시기~번데기 시기를 알곡 안에서 지나기 때문에, 이 알곡의 크기에 규제되어 몸의 크기를 변화시킨다. 앞에 인용한 들로벨 씨 등의 실험에서는 메밀, 타마린드, 완두콩, 밤, 도토리 등의 곡물과는 다른 먹을거리에서 어리쌀바구미, 바구미, 적갈색쌀바구미를 키웠는데, 밤이나 도토리에서 사육한 바구미의 중량이 가장 크고, 난소 안의 공생 박테리아 수도 가장 많았다. 중량이 크다고 하는 것은 몸도 크다고 하는 것을 의미한다. 이 높은 발육에는 밤이나 도토리에 포함된 당분이 크게 작용할 가능성이 지적된다(Delobel and Grenier 1993).
필자 등에 의한 생육실험에서는 중량이 아닌 겉날개나 흉부의 길이나 너비 등을 이용해 비교한다. 그 경우, 도토리나 밤에서 사육한 어리쌀바구미는 쌀알에서 키운 어리쌀바구미보다 30% 정도 커지는 것이 판명된다. 사실 압흔 어리쌀바구미도 벼에서 사육된 현생 어리쌀바구미보다 30% 정도 크다(그림30).
그들의 선조가 본래의 생태, 생육 실험의 결과 예상되는 죠몬 시대의 건조 저장 가능한 먹을거리 등을 종합적으로 판단하면 그 가해 대상물은 밤이나 도토리였다고 생각할 수 있다. 그외에 대두나 팥, 또는 조릿대의 열매 등 건조 보존이 가능한 먹을거리도 그들의 가해 대상이 되었을 가능성이 있다.
정주 마을과 저장 먹을거리
현재, 전국에 51개소의 유적에서 어리쌀바구미 압흔이 검출된다. 아직 사례가 적기에 결론을 내는 것은 성급할지도 모르지만, 지금까지 어리쌀바구미 압흔이 검출되는 유적는 그 규모에서 보아 정주성이 높은 마을 유적이 많다는 인상이 있다. 가고시마현 산본마츠 유적의 경우도 죠몬 시대 조기 전반이라고는 하나 평평한 바닥의 원통 토기를 지닌 수혈 주거터가 있는 정주 성격의 마을이다. 또한 다량의 어리쌀바구미 압흔을 검출한 미야자키현 모토노바루本野原 유적도 큐슈 최초의 본격적인 환상 마을이고, 산본마츠 유적에 다음가는 오래된 어리쌀바구미 압흔을 검출한 죠몬 시대 조기 후반의 사가현佐賀県 히가시묘우東名 유적도 온난화로 인해 정주성이 증가하는 시기의 유적이다. 또한 도호쿠 지방의 산나이마루야마 유적도 호쿠리쿠 지방의 히라오카平岡 유적이나 사쿠라마치桜町 유적도 그 땅의 정주적인 큰 마을이다. 앞에 예측했듯이, 압흔 어리쌀바구미가 실내 해충, 더구나 저장 먹을거리 해충이었다고 한다면, 그와 같은 벌레가 꾀기 위해서는 먹을거리의 저장 기간이 몇 개월 단위일 필요가 있기에, 그들의 존재 자체가 장기 거주의 증거가 된다.
또한 유적의 분포를 보면, 따뜻한 지방만이 아니라, 한랭한 도호쿠 지방의 아오모리현 산나이마루야마 유적이나 호쿠리쿠 지방의 도야마현 히라오카 유적이나 같은 사쿠라마치 유적에서도 발견된다. 모두 죠몬 시대 전기라는 비교적 따뜻했던 시기라고는 하나, 한랭한 지역에서 적응한 건 인위적 환경이 아니면 일어날 수 없다. 현생 바구미의 일본 열도 분포를 보면, 월 평균기온 17도 이하의 지역에서는 매우 분포밀도가 낮아진다. 다만, 추운 지역이더라도 따뜻한 짧은 여름이 있다면 충분히 번식은 가능하여, 거점적인 마을에서 1년 중 그 약 절반 정도의 시기, 특히 초여름부터 초가을에 걸쳐 점령지가 되는듯한 마을에서는 어리쌀바구미는 번식할 수 있었을 것이라 생각할 수 있다.
사람이 날랐다?
그럼 그들이 어떻게 확산해 나아갔을까? 지금이야 대량 무역의 시대이고, 세계적인 저곡 해충의 만연이 상징하듯이 그들은 식량 운반과 저장장 창출의 반복에 의하여 세계로 확산된다. 유럽에서 적갈색쌀바구미의 확산은 신석기 시대의 농경 전파의 증거로도 다루어지는데, 이후 로마군에 의한 병량의 운반과 비축에 의하여 더 빠르고 더욱 광대한 지역으로 확산되었단 것이 밝혀졌다. 앞에 기술했듯이, 어리쌀바구미속 갑충은 인위적인 운반이 없으면 원거리 이동, 정착은 할 수 없다. 현재, 어리쌀바구미 압흔은 나가사키현의 고토우五島, 가고시마현의 타네가시마種子島, 아마미오오시마奄美大島, 도쿠노시마徳之島, 오키나와현의 이에지마伊江島 등 도서부의 유적에서도 다수 검출된다. 이는 각각의 섬에 서식하고 있던 재래종이 죠몬인들의 도토리 저장에 대하여 각지에서 똑같이 적응했다고 생각하기보다, 식량과 함께 운반되어 섬을 따라 확산된 결과라고 생각하는 쪽이 이해하기 쉽다. 유럽의 연구에서도 적갈색쌀바구미는 추위에 강하지만, 북유럽 지역으로 확산되는 데에는 인위적 운반이 필요했다고 생각된다. 추운 지역으로 확산됨과 바다를 넘어 확산되는 데에는 인간에 의한 가해된 식량의 운반, 가지고 감이 배경에 있다고 상정되는 것이다.
어리쌀바구미 압흔이 찍히는 방법
어리쌀바구미가 도토리나 밤을 비롯한 정주 마을의 건조 먹을거리의 저장소에 생식했던 것은 확정할 수 있었다. 그러면 왜, 어떻게 하여 압흔으로 남게 된 것일까?
토기 압흔이 찍히는 방법은 곤충이 토기 제작 과정 중에 언제 토기의 점토 안에 들어갔는지를 추정하는 수단이 된다(표3). 어리쌀바구미 성충 298점의 압흔이 찍힌 방법을 조사해 보면, 토기를 제작할 때 토기 위를 보행하다가 눌려서 점토 안에 들어갔을 가능성이 있는 상태는 토기의 안팎 벽면이라면 어리쌀바구미의 배 쪽, 토기 바닥면이라면 등 쪽이 압흔으로 남을 터이다(표에서 굵은선 안). 그와 같은 상태는 40%도 되지 않고, 나머지는 등 또는 옆 쪽의 압흔으로 그 방향성은 일정하다고 할 수 없다. 또한 결손율은 7.3%이고, 개체 중에는 흉부나 복부만 있는 것이나 전체 모양은 남았지만 두부가 부러져 뒤로 꺾인 것 등 상당한 손상을 입은 것을 알 수 있다. 나아가 4가지 예는 단면으로 검출된 예로, 이들은 본래 토기 표면에 표출되지 않는 '잠재 압흔'이고, 토기가 깨져서 단면으로 노출된 것이다. 또한 최근 도야마현 히라오카 유적(죠몬 시대 전기 후엽), 미야자키현 모토노바루 유적(죠몬 시대 후기 전엽), 가고시마현 츠카가단塚ヶ段 유적(죠몬 시대 후기 말), 가고시마현 코자코小迫 유적(죠몬 시대 만기 말), 가고시마현 고쿠라마에 유적(돌대문 시기)에서 연엑스선, 엑스선 CT에 의하여 어리쌀바구미의 잠재 압흔을 검출한다. 이와 같은 점에서 보아, 곤충 압흔의 대부분은 토기를 제작할 때 토기 표면을 기어가던 것이 눌려 들어간 압흔(impression)이 아니라, 토기 바탕흙 속에 반죽되어 들어간 껍데기(cavity), 즉 바탕흙에 들어간 '혼입체의 흔적'이 주체를 차지하고 있다는 것을 알 수 있다.
| 어리쌀바구미의 방향 | 토기의 검출 부위 | 엑스선 CT | |||
| 주둥이, 몸통, 안바닥 | 바깥바닥 | 단면 | |||
| 등 쪽 | 73(24.6%) | 9(3.0%) | 4(1.3%) | 6(2.0%) | |
| 옆 쪽 | 98(33.0%) | 4(1.3%) | |||
| 배 쪽 | 101(34.0%) | 3(1.0%) | 총계 | ||
| 합계 | 272(91.6%) | 16(5.4%) | 4(1.3%) | 6(2.0%) | 298(100.0%) |
압흔 형성의 시기
어리쌀바구미가 바탕흙에 혼입될 기회는 앞의 압흔 산출 상태 분석의 결과에서 보면, 바탕흙에 혼화재로 식물섬유를 넣을 때, 또는 점토가래 만들기의 두 단계를 생각할 수 있다.
후자의 경우는 점토를 반죽하고 있을 때 우연히 그곳을 기어 가던 어리쌀바구미가 점토가래에 휘말려 들어가, 즉 우연의 혼입이다.
이에 반해 전자의 경우는 어느 정도 의도적이다. 적어도 죠몬인들은 어리쌀바구미의 존재는 의식하고 있었다고 생각된다. 혼화재에는 식물섬유 이외에 해충에 가해된 도토리나 밤 등의 전분질 건조 먹을거리의 가루 등도 상정할 수 있다. 토기 형성에 필요한 적당한 점토의 가소성을 얻는 데에는 '재우는' 과정에서 일어나는 박테리아에 의한, 어느 종의 발효가 필요하고, 도자기의 경우이긴 하지만 단기간에 '재우는' 효과를 높이기(박테리아의 증가를 촉진함) 위하여 전분이나 설탕 같은 유기질을 소량 첨가하는 예가 소개되어 있다(後藤 1980). 유럽 신석기 시대의 맥류 저장 해충인 적갈색쌀바구미가 토기 압흔으로 검출되는 예가 두 가지 정도 소개되어 있다(Buckland 1990, Panagiotakopulu 2010). 이 한 가지는 보리의 나락과 함께 혼입되고, 혼화재에 섞여서 저곡 해충이 들어갔을 가능성을 보여주는 좋은 예라 할 수 있다.
마찬가지로 토기의 개체에 한 종류의 곤충이 다수 혼입된 토기의 존재를 생각하면, 더 적극적으로 곤충 자체를 의도적으로 혼입했을 가능성도 전혀 없을 리는 없다. 오히려 지금까지 발견된 콩류나 들깨 등의 식물 씨앗의 압흔 형태로 보면, 의도적이었다고 하는 평가를 받을 수도 있다. 그러나 이것을 입증하기 위해서는 토기 안에서 발견되는 식물 씨앗의 종류, 그들의 껍질이나 부속품의 유무, 동물 섬유나 식물 섬유 등의 혼화재 넣는 방식 등을 상세히 검증해야 한다. 앞으로의 과제이다.
대륙에서 발견되길 기대함
산본마츠 유적의 어리쌀바구미 압흔은 약 1만 년 전에 큐슈 본섬에 근접한 타네가시마에서 어리쌀바구미가 이미 존재한 것을 보여주고, 그 존재는 농경(벼농사) 확산의 이론으로는 설명할 수 없는 것, 그리고 자매종이라 하는 적갈색쌀바구미, 바구미를 합한 3종의 분화와 기원지에서의 확산이 지금까지 상정된 시기보다 오랜 시기에 이미 일어났던 것을 보여준다.
조선반도나 중국 대륙에서는 원래 유적에서 발견되는 어리쌀바구미속 갑충 자체가 적은 데다, 산본마츠 유적에 필적할 만한 오랜 시기의 신석기 시대의 어리쌀바구미 발견은 없고, 확산 시기가 언제였는지, 충분한 검증을 할 수 없는 상황에 있다. 그러나 중국의 전한 시대에는 바구미가 동정되어 있고, 죠몬 시대는 거의 모든 것이 어리쌀바구미라고 생각되는 점에서 어리쌀바구미는 일본 원생의 것일 가능성도 있다.
고고 유적은 화석 자료와 현생 어리쌀바구미 사이를 매우고 해충으로서 진화하는 과정을 해명할 열쇠를 쥐고 있다. 적어도 말할 수 있는 건, 일본 열도에서는 1만 년 전 이후 인간이 도토리나 밤을 채집해 그것들을 축적하게 되어 어리쌀바구미가 기생, 해충화했다. 현재의 어리쌀바구미는 지금으로부터 3000년 전 무렵 벼의 유입을 계기로 벼에 특화해 소형화한 것일까, 아니면 이미 벼에 기생하던 작은 어리쌀바구미가 전래되어 재래 도토리 어리쌀바구미와 융합한 것일까? 이 수수께끼를 푸는 열쇠는 역시 어리쌀바구미의 크기에 있다.
도서부에서 발견되는 어리쌀바구미 압흔 가운데 가고시마현 도쿠노시마의 오모나와面縄 제1패총(카네쿠식兼久式 토기)이나 오키나와현 이에지마의 나가라바루히가시ナガラ原東 유적(고분 시대 상당 시기)의 어리쌀바구미 압흔은 죠몬 시대의 어리쌀바구미 압흔에 비하여 특별히 작은 건 아니다. 이 지역에서 벼농사의 전파는 구스쿠グスク 시대이며, 그 이전의 어리쌀바구미는 시대가 새로워져도 견과류에 적응해 이어진 자손이었다고 생각할 수 있다. 마찬가지로, 일본 혼슈 동쪽의 야요이 시대 전기 상당 시기의 예도 벼에 특화된 특별히 작은 크기는 아니다. 따라서 일본의 어리쌀바구미에게 벼에 대한 특화(쌀을 먹는 벌레의 등장)는 야요이 시대 중기 이후 또는 그보다 훨씬 뒤였을 가능성도 있다. 이와 같은 추론을 증명하기 위해서도 국내 유적에 대한 추가적인 조사와 함께, 중국이나 조선반도, 동남아시아 여러 나라의 어리쌀바구미속 갑충의 고고학 자료의 발굴이 필요할 것이다. 의식적인 조사가 요망된다.
죠몬인의 어리쌀바구미에 대한 추억
2012년 9월 30일에 키시와다시岸和田市 시립문화회관에서 개최된 하마다 세이료우濱田青陵 상 수상기념 심포지엄에서 어리쌀바구미가 왜 토기 점토 안에 들어갔는지 하는 사회자 아마노 유키히로天野幸広 씨의 질문에 나는 "맛있는 도토리나 밤을 먹고 많은 자손을 남기는 벌레들을 보고 죠몬인들도 그걸 닮고 싶다고 하며 번영을 기리며 넣은 것은 아닐까요?"라고 답했다. 그러나 아마노 씨는 "도토리 등의 귀중한 식량을 가해하는 어리쌀바구미가 미워서 토기 안에 봉해 버린 것은 아닐까요?"라는 의견을 내놓았다. 정면으로 대립되는 의견이다. 나는 자주 강연회 등에서 내가 마을의 유적에 해충이 있었다는 말을 하면 약간 싫은 기색을 내비치는 청중들에게 어리쌀바구미의 존재는 풍부한 저장 식량이 있었던 풍요와 번영의 반증이라 한다고 긍정적인 설명을 하기로 했다. 다만, 실제 해충은 프랑스 혁명의 방아쇠가 되었을 정도로 문명에 영향을 미치는 부정적인 유산이기도 하다. 확실히 중요하게 보존되던 먹을거리를 먹어치우던 원한은 컸을 것이다.
지금은 죠몬인들의 기분을 충분히 알 수 없지만, 이 어리쌀바구미 압흔을 철저하게 발굴해 나아가면, 머지않아 그들의 어리쌀바구미에 대한 생각을 밝힐 수 있을 것이라 확신한다. 작은 벌레이지만, 죠몬인의 살림살이나 정신성을 밝힐 수 있는 큰 잠재력을 내포한 연구자료라고 생각한다.
'농담 > 농-문화' 카테고리의 다른 글
| 씨앗을 심는 죠몬인 -압흔법이 밝힌 것 (0) | 2025.01.06 |
|---|---|
| 씨앗을 심는 죠몬인 -벼는 언제 일본에 전해졌을까? (0) | 2024.12.30 |
| 씨앗을 심는 죠몬인 -죠몬인은 풍족한 수렵채집민인가 (0) | 2024.10.22 |
| 일본 민가 집락 박물관 (0) | 2024.10.10 |
| 씨앗을 심는 죠몬인 -대두와 죠몬인 (0) | 2024.10.09 |



